Research Areas

Cognition
Cognition emerges as an interaction between top-down and bottom-up processing. Bottom-up processing carries sensory information about stimuli in the environment, such as color, form, shape, and contrast in vision. Top-down processing represents our internal thoughts, goals, attention, and predictions.
Bottom-up and top-down processing take place over a number of both cortical and subcortical processing stages. They involve computations within each stage and communication between stages. To understand these processing streams and how they are integrated, we need to understand the brain holistically. This involves recording neural activity in multiple brain regions simultaneously. Then, we can understand how many interacting parts (neurons within brain areas) can give rise to emergent, network-level phenomenon (cognition and consciousness).
A delayed match to sample task can model bottom-up and top-down processing in non-human primates (NHPs). NHPs are trained to visually fixate on a sample stimulus that appears at the center of the screen. The sample stimulus disappears during a short delay period and reemerges with two distractor stimuli. During the response period, NHPs saccade (movement of eye fixation) to the previously sampled stimulus, while ignoring the distractors. This can be measured through eye tracking technology. This task design provides a simple yet powerful way to model different aspects of cognition. The response period models attention and selection processes. The delay period models working memory. The sample window allows us to measure the effects of prediction on sensory responses. Through brain recordings, our laboratory can investigate the underlying neural mechanisms of these cognitive processes.

After a 1-s fixation window, a sample stimulus was shown for 1 s. After a delay, the target reappeared at one of four locations. NHPs saccaded to the sampled stimulus while ignoring the distractors. From Bastos et al. (2020)
An example of the delayed match to sample task our laboratory investigates in NHPs. The video displays a correct trial, showing the NHP fixating on the sample stimulus (rainbow), which disappears, then reappears with two additional distractors. The NHP saccades to the correct sampled stimulus and receives a tasty juice reward for this correct trial.
Deficits in Prediction, Attention, and Working Memory are at the core of many brain disorders. For example, autism spectrum disorders can be understood as a condition in which the brain operates with faulty predictions of social interaction. Every social interaction becomes “surprising” and thus overwhelming. Schizophrenia can be understood as a brain disorder where overly tight predictive models overrule sensory inputs and generate hallucinations. This causes Working Memory deficits and a loss of ordered control of thought. Depression can be understood as a strong prediction that future states will be negative, and a failure to attend to positive emotions. For these reasons, it is imperative that we gain a better understanding of the fundamental mechanisms by which cognition works in the brain. These insights will ultimately translate into a better understanding of human nature as well as novel treatments for these and other brain disorders.

Computation
How do networks of neurons process their inputs and transform them into more useful outputs? This is what we mean by neuronal computation. One example of a neuronal computation is selective attention: at a cocktail party, we can easily attend to the conversation we are having while ignoring other conversations that may be just as loud (that is, until we hear a juicy tidbit of gossip). What is going on here? Neurons that represent the attended information stream (the conversation we are focused on) enhance their firing rates, decrease their noise correlation, increase their gamma frequency (40-100 Hz) synchronization, and decrease lower frequency synchronization in the alpha/beta (8-30 Hz) frequency range and below. These excitatory and inhibitory network computations work together to enable our attention to stay focused.

When inhibitory and excitatory neurons wire together, they can generate different brain waves in different layers of cortex. Superficial layers are rich in fast gamma waves (depicted in blue) and deep layers are rich in slow alpha/beta waves (depicted in red). Alpha/beta in deep layers inhibits gamma in superficial layers. Over time, moving from left to right in the figure, the deep layer alpha/beta frequency decreases and releases inhibition in superficial layers, resulting in increased superficial gamma power and spiking (black ticks). From Bastos et al. (2018).
Emerging evidence suggests that neural computations rely on distinct layers of cortex, neuronal cell types, and synchronous brain rhythms from 1-100 Hz (also known as "brain waves"). The Bastos laboratory primarily works with non-human primates (NHPs) as they play simple videogames, which model cognitive tasks. As NHPs perform these tasks, we use large-scale multi-laminar recordings to pinpoint the exact brain regions, cortical layers, and neural oscillations used by the brain to solve the task. We use advanced data analysis techniques to mine these rich data for insights. We use these tools to decipher where and when computations are performed and how these computations are communicated between cortical regions.
As we acquire richer experimental data, we can also build computational models of the underlying circuitry. This can help us explore a much richer parameter space than we could ever measure empirically. These computational models inform us as to where our knowledge is missing, so we can design the crucial experiments to fill in the gaps. We also use techniques such as optogenetics and microstimulation to perturb ongoing neuronal activity and test which computations are critical for cognition. One such computational model is the Predictive Routing Model, in which superficial cortical layers (1 to 4) and deep cortical layers (5/6) form a computational network to prediction. Deep cortical layers are prepared to process a predicted stimulus from feedback communication in superficial cortical layers, increasing inhibitory alpha/beta synchronization and therefore inhibits gamma spiking. Consequently, superficial-layer processing of the predicted stimulus is inhibited by decreased excitatory gamma synchronization, reducing feedforward communication. When a strong prediction is made and something different occurs (a prediction error), there is less feedback alpha/beta inhibition, causing an excitatory response of increased gamma spiking and enhanced feedforward communication from deep layers to superficial layers that updates the prediction model to fit the current environment.
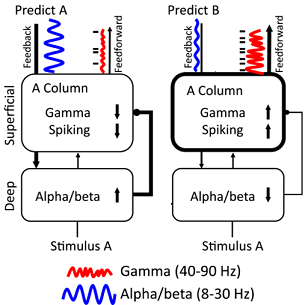
Predictive Routing Model. (Left subpanel) Feedback communication occurs through deep alpha/beta synchronization when predicting Stimulus A, inhibiting gamma spiking in superficial-layer processing, which reduces feedforward outputs. (Right subpanel) When a prediction (Predict B) is made and a prediction error occurs (Stimulus A), there is less feedback alpha/beta inhibition, causing the A column to respond to the surprising stimulus with more gamma/spiking and enhanced feedforward communication from superficial layers to update the predictive model. From Bastos et al. (2020).

Consciousness
How do these neuronal computations give rise not only to cognition, but also the experience of being conscious? Different anesthetic agents such as propofol or ketamine are used on a daily basis in medicine to temporarily render patients unconscious or to manipulate consciousness in therapeutic ways. In the Bastos Lab, our framework for understanding the neuronal basis of consciousness is to ask: which particular neuronal computations are taken offline during states of unconsciousness and altered consciousness?
We have recently discovered that throughout frontal, parietal, and temporal cortex, anesthetic drugs such as propofol induce loss of consciousness (LOC) by powerfully controlling the brain's oscillations. Propofol induces the emergence of a slow frequency (~ 1 Hz) oscillation that synchronizes these cortical areas together with a deeper brain structure, the thalamus. These slow frequency oscillations create long periods (of about 1 second) of near-silence when almost no neuronal spiking occurs. These “Off” periods are interspersed with “On” periods of activity. This is quite different from the normal state of wakefulness where brain activity is dominated by much higher frequency background activity. The profound shift in slow frequency dynamics during unconsciousness leads to a brain which is no longer capable of functionally integrating inputs across multiple areas. Bottom-up and top-down processing are unplugged and disconnected. The neuronal computations that lead to integration across processing stages and consciousness are offline. We also observed that thalamic microstimulation safely and effectively reversed these effects. Thalamic microstimulation broke apart the slow frequency synchronization and subjects regained arousal. This is an example of how using an anesthetic agent (propofol) can lead to insights that link neuronal activity across brain regions to consciousness.
We plan to study the effects of multiple other types of drugs on neuronal computations and cognition to gain insight into how these processes enable different conscious states.

Physiological changes during anesthesia induced LOC. (upper panel) Neural activity during the Awake (left) and Unconscious states (right) showing clear slow-frequency waves. (lower panel) Neuronal spiking times are indicated with dots, which displays the “off and on” periods of activity during LOC (right). From Bastos et al. (2021)